
Energía en seres vivos |
La energía en los seres vivos se obtiene mediante una molécula llamada ATP (adenosín trifosfato) .
Aunque son muy diversas las biomoléculas que contienen energía almacenada en sus enlaces, es el ATP la molécula que interviene en todas las transacciones (intercambios) de energía que se llevan a cabo en las células; por ella se la califica como "moneda universal de energía".
El ATP está formado por adenina, ribosa y tres grupos fosfatos, contiene enlaces de alta energía entre los grupos fosfato; al romperse dichos enlaces se libera la energía almacenada.
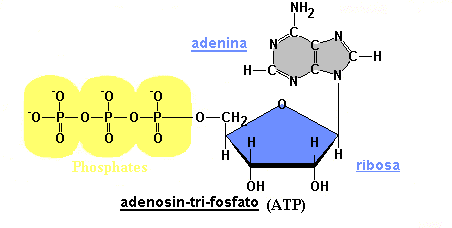
|
En la mayoría de las reacciones celulares el ATP se hidroliza a ADP (adenosín difosfato), rompiéndose un solo enlace y quedando un grupo fosfato libre, que suele transferirse a otra molécula en lo que se conoce como fosforilación; sólo en algunos casos se rompen los dos enlaces resultando AMP (adenosín monofosfato) + 2 grupos fosfato.
El sistema ATP <—-> ADP es el sistema universal de intercambio de energía en las células.
Los procesos celulares que llevan a la obtención de energía (medida en moléculas de ATP) son la fotosínteis y la respiración celular :
Fotosíntesis
La fotosíntesis es uno de los procesos metabólicos de los que se valen las células para obtener energía.
Es un proceso complejo, mediante el cual los seres vivos poseedores de clorofila y otros pigmentos captan energía luminosa procedente del sol y la transforman en energía química (ATP) y en compuestos reductores (NADPH), y con ellos transforman el agua y el CO 2 en compuestos orgánicos reducidos (glucosa y otros), liberando oxígeno :
|
CO
2
+ H
2
O+ LUZ
|
La energía captada en la fotosíntesis y el poder reductor adquirido en el proceso, hacen posible la reducción y la asimilación de los bioelementos necesarios, como nitrógeno y azufre, además de carbono, para formar materia viva .
La radiación luminosa llega a la tierra en forma de "pequeños paquetes", conocidos como cuantos o fotones . Los seres fotosintéticos captan la luz mediante diversos pigmentos fotosensibles, entre los que destacan por su abundancia las clorofilas y carotenos.
Al absorber los pigmentos la luz, electrones de sus moléculas adquieren niveles energéticos superiores, cuando vuelven a su nivel inicial liberan la energía que sirve para activar una reacción química: una molécula de pigmento se oxida al perder un electrón que es recogido por otra sustancia, que se reduce . Así la clorofila puede transformar la energía luminosa en energía química.
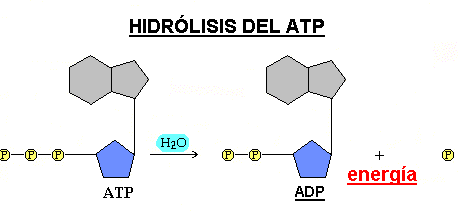
|
En la fotosíntesis se diferencian dos etapas, con dos tipos de reacciones:
1) Fase luminosa: en el tilacoide en ella se producen transferencias de electrones. Los hechos que ocurren en la fase luminosa de la fotosíntesis se pueden resumir en estos puntos:
a) Síntesis de ATP o fotofosforilación que puede ser:
acíclica o abierta
cíclica o cerrada
b) Síntesis de poder reductor NADPH
2) Fotólisis del agua
Los pigmentos presentes en los tilacoides de los cloroplastos se encuentran organizados en fotosistemas (conjuntos funcionales formados por más de 200 moléculas de pigmentos); la luz captada en ellos por pigmentos que hacen de antena , es llevada hasta la molécula de "clorofila diana" que es la molécula que se oxida al liberar un electrón, que es el que irá pasando por una serie de transportadores, en cuyo recorrido liberará la energía .
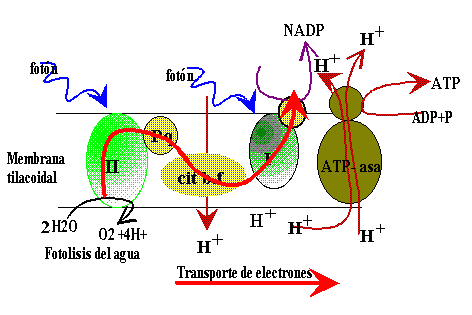
|
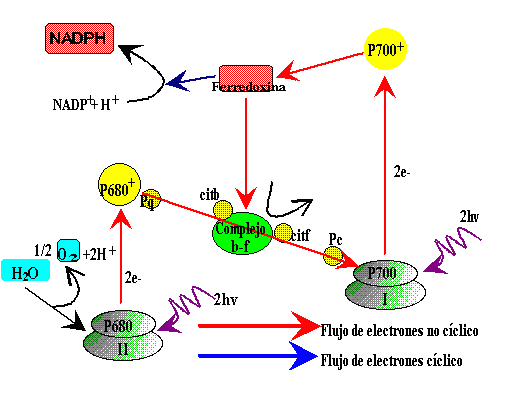
Existen dos tipos de fotosistemas, el fotosistema I (FSI), está asociado a moléculas de clorofila que absorben a longitudes de ondas largas (700 nm)y se conoce como P700. El fotosistema II (FSII), está asociado a moléculas de clorofila que absorben a 680 nm. por eso se denomina P680.
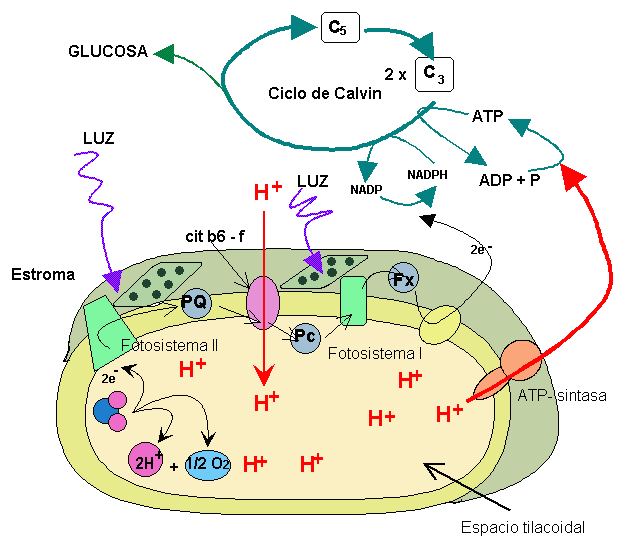
La luz es recibida en el FSII por la clorofila P680 que se oxida al liberar un electrón que asciende a un nivel superior de energía; ese electrón es recogido por una sustancia aceptor de electrones que se reduce, la Plastoquinona (PQ) y desde ésta va pasando a lo largo de una cadena transportadora de electrones, entre los que están varios citocromos (cyt b/f) y así llega hasta la plastocianina (PC) que se los cederá a moléculas de clorofila del FSI.
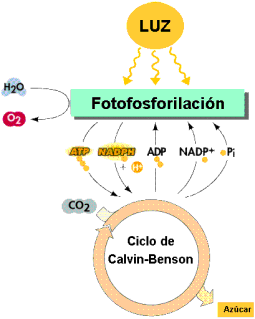
En el descenso por esta cadena, con oxidación y reducción en cada paso, el electrón va liberando la energía que tenía en exceso; energía que se utiliza para bombear protones de hidrógeno desde el estroma hasta el interior de los tilacoides, generando un gradiente electroquímico de protones . Estos protones vuelven al estroma a través de la ATP-asa y se originan moléculas de ATP.
El fotosistema II se reduce al recibir electrones procedentes de una molécula de H2O, que también por acción de la luz, se descompone en hidrógeno y oxígeno, en el proceso llamado fotólisis del H2O. De este modo se puede mantener un flujo continuo de electrones desde el agua hacia el fotosistema II y de éste al fotosistema I.
|
En el fotosistema I la luz produce el mismo efecto sobre la clorofila P700, de modo que algún electrón adquiere un nivel energético superior y abandona la molécula, es recogido por otro aceptor de electrones, la ferredoxina y pasa por una nueva cadena de transporte hasta llegar a una molécula de NADP+ que es reducida a NADPH, al recibir dos electrones y un protón H + que también procede de la descomposición del H 2 O.
Los dos fotosistemas pueden actuar conjuntamente –proceso conocido como esquema en Z – para producir la fotofosforilación (obtención de ATP) o hacerlo solamente el fotosistema I; se diferencia entonces entre fosforilación no cíclica o acíclica cuando actúan los dos, y fotofosforilación cíclica , cuando actúa el fotosistema I únicamente. En la fotofosforilación acíclica se obtiene ATP y se reduce el NADP+ a NADPH , mientras que en la fotofosforilación cíclica únicamente se obtiene ATP y no se libera oxígeno.
Mientras la luz llega a los fotosistemas, se mantiene un flujo de electrones desde el agua al fotosistema II, de éste al fotosistema I, hasta llegar el NADP + que los recoge; ésta pequeña corriente eléctrica es la que mantiene el ciclo de la vida.
Fase oscura: en el estroma . En ella se realiza la fijación de carbono. En esta fase, se va a utilizar la energía química obtenida en la fase luminosa, en reducir CO 2 , Nitratos y Sulfatos y asimilar los bioelementos C, H, y S, con el fin de sintetizar glúcidos, aminoácidos y otras sustancias.
|
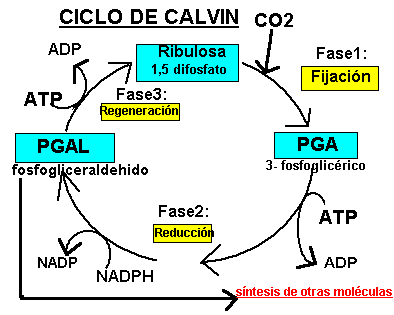
Las plantas obtienen el CO 2 del aire a través de los estomas de sus hojas. El proceso de reducción del carbono es cíclico y se conoce como Ciclo de Calvin , en honor de su descubridor M. Calvin.
La fijación del CO2 se produce en tres fases:
Carboxilativa : El CO2 se fija a una molécula de 5C, la ribulosa 1,5 difosfato , formándose un compuesto inestable de 6C, que se divide en dos moléculas de ácido 3 fosfoglicérico conocido también con las siglas de PGA.
Reductiva: El ácido 3 fosfoglicérico se reduce a gliceraldehido 3 fosfato , también conocido como PGAL ,utilizándose ATP Y NADPH.
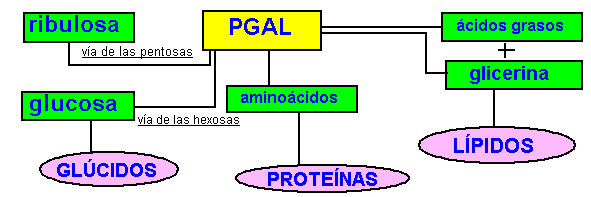
3.Regenerativa/Sintética: Las moléculas de gliceraldehido 3 fosfato formadas siguen diversas rutas; de cada seis moléculas, cinco se utilizan para regenerar la ribulosa 1,5 difosfato y hacer que el ciclo de calvin pueda seguir, y una será empleada para poder sintetizar moléculas de glucosa (vía de las hexosas), ácidos grasos , aminoácidos ... etc; y en general todas las moléculas que necesita la célula.
|
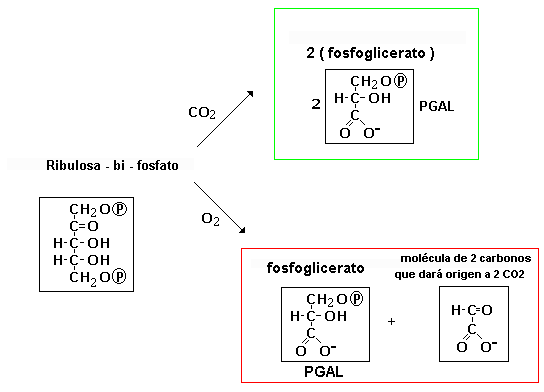
En el ciclo para fijar el CO 2 , intervienen una serie de enzimas, y la más conocida es la enzima Rubisco ( ribulosa 1,5 difosfato carboxilasa/oxidasa ), que puede actuar como carboxilasa o como oxidasa, según la concentración de CO 2 .
|
|
Si la concentración de CO 2 es baja, funciona como oxidasa, y en lugar de ayudar a la fijación de CO 2 mediante el ciclo de Calvin, se produce la oxidación de glúcidos hasta CO 2 y H2O, y al proceso se le conoce como fotorrespiración. La fotorrespiración no debe confundirse con la respiración mitocondrial, la energía se pierde y no se produce ni ATP ni NADPH; y como se ve en el esquema se disminuye el rendimiento de la fotosíntesis, porque sólo se produce una molécula de PGA que pasará al ciclo de Calvin; en cambio cuando funciona como carboxilasa, se obtienen dos moléculas de PGA.
Hipótesis quimiosmótica de la fotofosforilación
La síntesis de ATP en el cloroplasto se explica mediante la hipótesis quimiosmótica de Mitchell, de forma muy semejante como ocurre en la mitocondria. El transporte de electrones en la cadena transportadora de la membrana tilacoidal produce el bombeo de protones desde el estroma hacia el espacio tilacoidal a nivel del complejo citocromo b6 - f , lo que genera un gradiente electroquímico. El flujo de protones a favor del gradiente desde el espacio tilacoidal hasta el estroma, a través del canal de protones de la ATP - sintasa , activa la síntesis de ATP a partir de ADP y fosfato.
Los electrones se emplean para reducir el NADP + a NADPH. El ATP y el NADPH producidos de esta forma pueden utilizarse en la fase oscura para las reacciones de síntesis, en las que se reducen moléculas sencillas, como el CO 2 , para formar glúcidos .
|
Importancia biológica de la fotosíntesis
La fotosíntesis es seguramente el proceso bioquímico más importante de la Biosfera por varios motivos:1. La síntesis de materia orgánica a partir de la inorgánica se realiza fundamentalmente mediante la fotosíntesis; luego irá pasando de unos seres vivos a otros mediante las cadenas tróficas, para ser transformada en materia propia por los diferentes seres vivos.
2. Produce la transformación de la energía luminosa en energía química , necesaria y utilizada por los seres vivos
3.En la fotosíntesis se libera oxígeno , que será utilizado en la respiración aerobia como oxidante.
4. La fotosíntesis fue causante del cambio producido en la atmósfera primitiva , que era anaerobia y reductora.
5. De la fotosíntesis depende también la energía almacenada en combustibles fósiles como carbón, petróleo y gas natural.
6. El equilibrio necesario entre seres autótrofos y heterótrofos no sería posible sin la fotosíntesis.
Se puede concluir que la diversidad de la vida existente en la Tierra depende principalmente de la fotosíntesis.
|
|
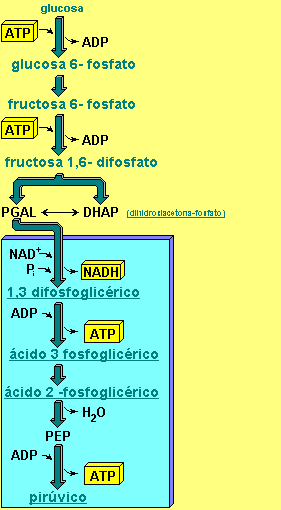
La glucólisis tiene lugar en el citoplasma celular. Consiste en una serie de diez reacciones, cada una catalizada por una enzima determinada, que permite transformar una molécula de glucosa en dos moléculas de un compuesto de tres carbonos, el ácido pirúvico.
En la primera parte se necesita energía, que es suministrada por dos moléculas de ATP, que servirán para fosforilar la glucosa y la fructosa . Al final de esta fase se obtienen, en la práctica dos moléculas de PGAL, ya que la molécula de DHAP (dihidroxiacetona-fosfato), se transforma en PGAL.
En la segunda fase, que afecta a las dos moléculas de PGAL, se forman cuatro moléculas de ATP y dos moléculas de NADH. Se produce una ganancia neta de dos moléculas de ATP .
Al final del proceso la molécula de glucosa queda transformada en dos moléculas de ácido pirúvico , es en estas moléculas donde se encuentra en estos momentos la mayor parte de la energía contenida en la glucosa.
La glucólisis se produce en la mayoría de las células vivas, tanto en procariontes como en las eucariontes.
|
Ciclo de Krebs
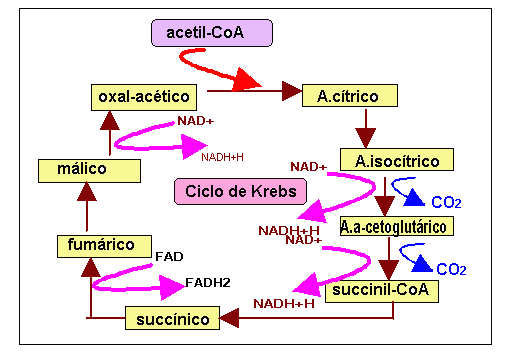
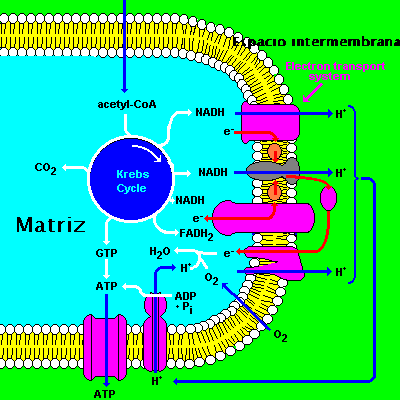
El producto más importante de la degradación de los carburantes metabólicos es el acetil-CoA, (ácido acético activado con la coenzima A), que continúa su proceso de oxidación hasta convertirse en CO 2 y H2O, mediante un conjunto de reacciones que constituyen el ciclo de Krebs punto central donde confluyen todas las rutas catabólicas de la respiración aerobia. Este ciclo se realiza en la matriz de la mitocondria
En este ciclo se consigue la oxidación total de los dos átomos de carbono del resto acetilo, que se eliminan en forma de CO 2 ; los electrones de alta energía obtenidos en las sucesivas oxidaciones se utilizan para formar NADH Y FADH2, que luego entrarán en la cadena respiratoria.
|
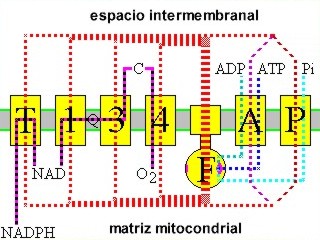
Cadena respiratoria
Sería la etapa final del proceso de la respiración celular, es entonces cuando los electrones "arrancados" a las moléculas que se respiran y que se "almacenan" en el NADH Y FADH2, irán pasando por una serie de transportadores, situados en las crestas mitocondriales formando tres grandes complejos enzimáticos.
La disposición de los transportadores permite que los electrones "salten" de unos a otros, liberándose una cierta cantidad de energía (son reacciones redox) que sirve para formar un enlace de alta energía entre el ADP y el P, que da lugar a una molécula de ATP.
El último aceptor de electrones es el oxígeno molecular y otra consecuencia será la formación de agua.
|
Hipótesis quimiosmótica
Según la hipótesis quimiosmótica sostenida por el investigador P. Mitchell, que es la que goza de mayor prestigio, y puede además explicar la síntesis de ATP tanto en la mitocondria como en el cloroplasto, la energía liberada por el transporte de electrones se utiliza para bombear protones desde la matriz al espacio intermembrana (en mitocondrias); o desde el estroma al interior del tilacoide (en cloroplastos). El bombeo de protones se realiza a través de transportadores localizados en complejos enzimáticas existentes en la membrana (de las crestas mitocondriales o membrana tilacoidal, según el caso).
De esta manera se genera un gradiente electroquímico de protones que ejerce lo que se conoce como fuerza protonmotriz , ya que cuando los protones atraviesan de nuevo la membrana interna (mitacondrial o tilacoidal) a favor del gradiente, lo hacen a través del sistema ATP-sintetasa , que se encuentra en dichas membranas, donde la energía protonmotriz se transforma en energía de enlace en moléculas de ATP .
El proceso se podría comparar con este símil:
El flujo de protones cumple el papel de transductor de energía, del mismo modo que el vapor que suministra una caldera puede utilizarse para generar energía eléctrica: el calor aplicado a la caldera (flujo de electrones) calienta el agua y forma vapor de agua (gradiente electroquímico de H+), cuya presión (fuerza protonmotriz) se puede acoplar a las turbinas de un generador eléctrico (ATP sintetasa) para producir electricidad (ATP).
|